У еукаріотичних клітинах основний обсяг генетичної інформації зосереджений у ядрі, де ДНК упакована в хромосоми разом з гістонами. Проте частина спадкового матеріалу розташована за межами ядерної оболонки — у цитоплазмі. Ця позаядерна ДНК, або цитоплазматична ДНК, міститься в спеціалізованих органелах, які походять від давніх бактерій. Питання про те, яка органела містить позаядерну ДНК, приводить до мітохондрій, що присутні майже в усіх еукаріотах, та хлоропластів у фотосинтезуючих організмах.
Мітохондрії та хлоропласти зберігають власні кільцеві молекули ДНК, які за будовою та механізмами реплікації нагадують бактеріальні хромосоми. Ці геноми забезпечують синтез обмеженої, але критично важливої кількості білків, необхідних для роботи органел. Решта білків, потрібних для функцій мітохондрій і хлоропластів, кодується ядерним геномом і імпортується з цитозолю. Така подвійна система контролю відображає давню еволюційну історію цих структур.
Вивчення позаядерної ДНК має практичне значення для медицини, сільського господарства та еволюційної біології. Мутації в мітохондріальній ДНК пов’язані з низкою спадкових захворювань людини, а хлоропластна ДНК використовується для ідентифікації видів рослин і створення трансгенних форм. Розуміння механізмів роботи цих геномів допомагає пояснити, чому деякі ознаки успадковуються не за менделівськими законами, а через цитоплазму.
Мітохондрії як універсальний носій позаядерної ДНК
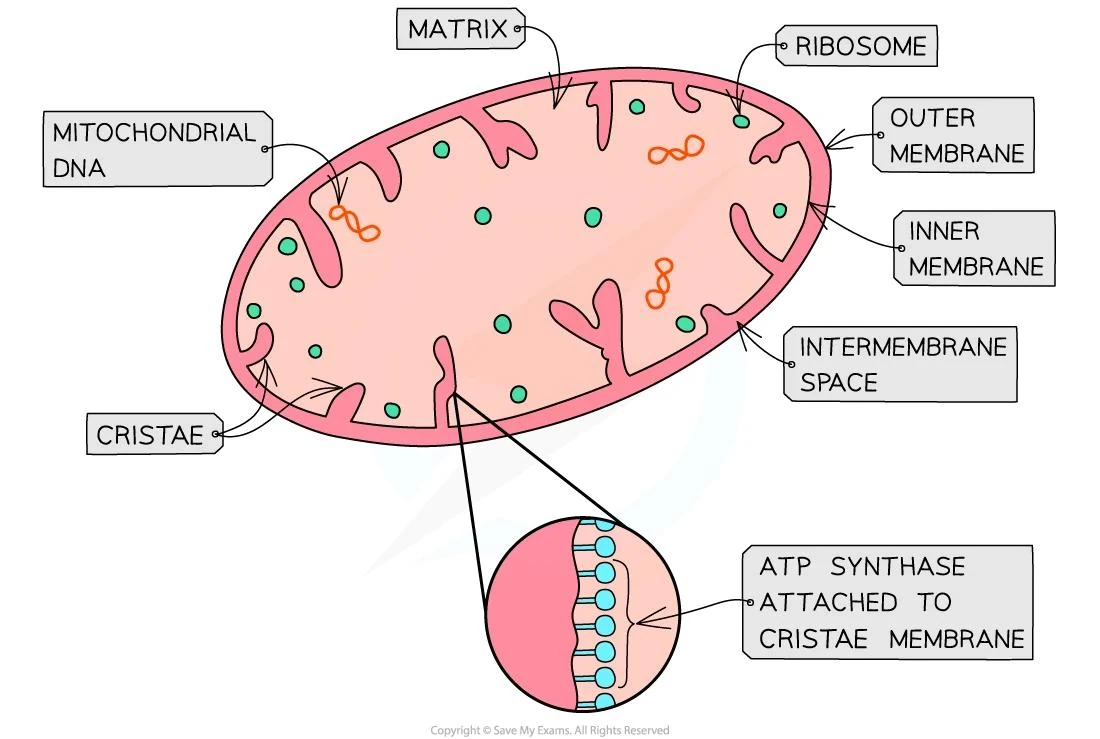
Мітохондрії — це двомембранні органели, що виконують роль енергетичних станцій клітини. Вони окислюють органічні речовини та синтезують АТФ через окисне фосфорилювання. У матриксі мітохондрій розташована мітохондріальна ДНК — компактна кільцева молекула, яка не пов’язана з гістонами. У людини така молекула має довжину 16 569 пар нуклеотидів і кодує 37 генів: 13 білків, 22 транспортні РНК та 2 рибосомні РНК.
Білки, що кодуються мітохондріальною ДНК, входять до складу комплексів електронно-транспортного ланцюга — I, III, IV та V. Ці субодиниці безпосередньо беруть участь у перенесенні електронів і синтезі АТФ. Інші компоненти цих комплексів, а також ферменти реплікації та транскрипції мітохондріальної ДНК, синтезуються в цитозолі за інформацією ядерних генів і транспортуються всередину органели. Така координація забезпечує ефективну роботу енергетичної системи клітини.
Мітохондріальна ДНК реплікується незалежно від клітинного циклу, хоча процес частково регулюється ядром. Молекули ДНК упаковані в нуклеоїди — компактні структури, що містять білок TFAM та інші фактори. В одній мітохондрії зазвичай присутні кілька копій геному, а в клітині їх може бути від сотень до тисяч. Успадкування мітохондріальної ДНК відбувається переважно по материнській лінії, оскільки яйцеклітина передає значно більше органел, ніж сперматозоїд.
Хлоропласти та їхній автономний геном у рослинних клітинах
Хлоропласти — це органели рослин і водоростей, у яких відбувається фотосинтез. Вони також містять власну позаядерну ДНК — хлоропластну ДНК, або пластидну ДНК. Ця молекула зазвичай має форму кільця довжиною від 120 до 160 тисяч пар нуклеотидів у наземних рослин, хоча в деяких водоростей розмір може відрізнятися. Геном хлоропласта кодує приблизно 100–130 генів, серед яких гени фотосистем I і II, АТФ-синтази, Рубіско (велика субодиниця), а також компоненти апарату транскрипції та трансляції.
Хлоропластна ДНК організована подібно до мітохондріальної: вона не містить гістонів, має бактеріальний тип рибосом (70S) і реплікується в стромі органели. Багато генів хлоропласта кодують білки, безпосередньо залучені до світлових реакцій фотосинтезу та фіксації вуглецю. Решта необхідних білків синтезується в цитозолі за ядерними генами і імпортується через специфічні транспортні системи оболонок хлоропласта.
У рослинній клітині хлоропласти успадковуються переважно від материнської рослини через яйцеклітину. Ця особливість використовується в селекції для закріплення певних ознак, наприклад, цитоплазматичної чоловічої стерильності, що полегшує отримання гібридного насіння. Хлоропластна ДНК також слугує зручним маркером для молекулярної ідентифікації видів рослин через свою консервативність і наявність варіабельних ділянок.
Порівняння геномів мітохондрій, хлоропластів та ядра
| Характеристика | Мітохондріальна ДНК | Хлоропластна ДНК | Ядерна ДНК |
|---|---|---|---|
| Розмір геному (приблизно) | 16,6 тис. пар нуклеотидів (людина) | 120–160 тис. пар нуклеотидів (наземні рослини) | Мільярди пар нуклеотидів |
| Форма молекули | Кільцева (переважно) | Кільцева (часто з інвертованими повторами) | Лінійні хромосоми |
| Кількість генів | 37 (людина) | 100–130 | 20–25 тисяч (людина) |
| Упаковка | Нуклеоїди, без гістонів | Нуклеоїди, без гістонів | Хроматин з гістонами |
| Рибосоми | 70S-подібні | 70S | 80S |
| Успадкування | Переважно материнське | Переважно материнське | Менделівське (батьківське + материнське) |
Дані узагальнено на основі публікацій ncbi.nlm.nih.gov. Таблиця демонструє ключові відмінності між трьома типами геномів еукаріотичної клітини. Позаядерні геноми значно менші за ядерний, але їхні продукти критично важливі для основних енергетичних процесів.
Ендосимбіотичне походження органел з позаядерною ДНК
Мітохондрії та хлоропласти вважаються нащадками вільноживучих бактерій, які були поглинуті давньою еукаріотичною клітиною. Згідно з ендосимбіотичною теорією, предком мітохондрій була аеробна альфа-протеобактерія, а предком хлоропластів — ціанобактерія. Свідченнями такого походження слугують кільцева форма ДНК, відсутність гістонів, наявність 70S-рибосом, здатність до автономного поділу шляхом бінарного поділу та послідовності генів, подібні до бактеріальних.
За мільярди років еволюції значна частина генів органел була перенесена до ядра. Цей процес ендосимбіотичного переносу генів пояснює, чому сучасні мітохондрії та хлоропласти залежать від ядерної інформації. Проте кілька десятків генів залишилися в органелах. Одна з гіпотез пов’язує це з необхідністю швидкого регулювання роботи електронно-транспортних ланцюгів безпосередньо на місці — у мембранах органел.
У деяких організмів, наприклад у паразитичних найпростіших, мітохондрії редукувалися до мітоzom або гідрогеносом, які іноді втрачають власну ДНК. Це демонструє пластичність еволюційного шляху органел, але в більшості еукаріотів мітохондрії зберігають функціональний геном.
Взаємодія позаядерних геномів з ядром та практичне значення
Робота мітохондрій і хлоропластів вимагає точної координації між ядерними та органельними генами. Ядро контролює більшість структурних і регуляторних білків, тоді як органели синтезують ключові компоненти енергетичних ланцюгів. Порушення цієї взаємодії призводить до накопичення мутацій у мітохондріальній ДНК, що пов’язано з віковими змінами, нейродегенеративними захворюваннями та онкологічними процесами.
У клінічній практиці аналіз мітохондріальної ДНК використовується для діагностики спадкових мітохондріальних хвороб, таких як синдром Лебера або MELAS. У судовій медицині та популяційній генетиці мітохондріальна ДНК служить маркером материнської лінії через відсутність рекомбінації та високу швидкість мутацій у певних ділянках.
У рослинництві хлоропластна ДНК застосовується для молекулярного маркування сортів і створення трансгенних рослин, у яких чужорідні гени інтегруються саме в пластидний геном. Це забезпечує високу експресію та материнське успадкування трансгену, що знижує ризик поширення через пилок.
Позаядерна ДНК залишається активною галуззю досліджень. Сучасні методи секвенування дозволяють вивчати гетероплазмію — присутність різних варіантів мітохондріальної ДНК в одній клітині — та її вплив на фенотип. Розуміння механізмів реплікації та репарації цих геномів відкриває перспективи для терапії мітохондріальних захворювань і вдосконалення біотехнологічних підходів у сільському господарстві.
Таким чином, відповідь на питання, яка органела містить позаядерну ДНК, охоплює мітохондрії як універсальний компонент еукаріотичних клітин та хлоропласти як специфічну особливість фотосинтезуючих організмів. Ці органели з власними геномами ілюструють складну еволюційну історію клітини та відіграють ключову роль у енергетичному забезпеченні життя.